








2023年(nián)10月(yuè)2日(rì),河南省農業科學院張海(hǎi)洋研究員(yuán)帶領的(de)芝麻基因組研究小(xiǎo)組(The Sesame
Genome Working Group)在《Plant
Communications》雜(zá)志(zhì)在線發表了題為(wèi)“Genomic evolution and insights into
agronomic trait innovations of Sesamum species”的(de)研究論文(wén)。該論文(wén)報道(dào)了胡麻屬芝麻栽培種和(hé)6個(gè)野生(shēng)種染色體(tǐ)級别的(de)基因組精細圖,首次揭示了胡麻屬物(wù)種的(de)進化(huà)規律及其在雙子(zǐ)葉植物(wù)進化(huà)中的(de)重要地(dì)位。研究發現(xiàn),在胡麻屬不同種群進化(huà)中,基因組結構變異對(duì)芝麻栽培種株型、含油量及枯萎病抗性水(shuǐ)平等重要農(nóng)藝性狀産生(shēng)了重大影響。發掘出了芝麻高(gāo)含油量基因SiNAC1(提高(gāo)6.2個(gè)百分點)、抗枯萎病基因ScaDIR40(免疫),首次揭示了芝麻栽培種較野生(shēng)種含油量高(gāo)而抗病性差的(de)機(jī)理(lǐ)。

芝麻(Sesamum indicum L.)屬胡麻科(kē)胡麻屬,起源于非洲,是世界上(shàng)最為(wèi)古老(lǎo)的(de)油料作(zuò)物(wù)之一。芝麻主要種植在熱帶和(hé)亞熱帶地(dì)區(qū),具有(yǒu)喜溫耐旱、耐瘠薄等優點,籽粒富含不飽和(hé)脂肪酸和(hé)抗氧化(huà)物(wù)質木酚素,被譽為(wèi)“油料皇後”。我國(guó)是世界芝麻四大主産國(guó)之一,也是最大的(de)消費(fèi)國(guó)。為(wèi)推動芝麻分子(zǐ)育種研究進程,2010年(nián)河南省農業科學院、南開(kāi)大學、河南工(gōng)業(yè)大學、江西(xī)省農(nóng)業(yè)科(kē)學院、安徽省農(nóng)業(yè)科(kē)學院等6家(jiā)芝麻科(kē)研團隊組建了芝麻基因組研究小(xiǎo)組,啓動了芝麻基因組計(jì)劃,并于2013年(nián)首次公布了芝麻栽培種De novo拼接基因組圖。
胡麻屬包含36個(gè)種(Species),涉及3個(gè)染色體(tǐ)組型(n = 13、16和(hé) 32),芝麻是胡麻屬中的(de)唯一栽培種。作(zuò)為(wèi)芝麻基因組計(jì)劃的(de)主體(tǐ)內(nèi)容,本研究構建了胡麻屬芝麻栽培種(S. indicum,n = 13)和(hé)6個(gè)野生(shēng)種(S. alatum,n=13; S. latifollum, n=16;S. angolense,n=16;S. calycinum,n=16;S. angustifolium,n=16;S. radiatum,n=32)染色體(tǐ)水(shuǐ)平的(de)的(de)基因組精細圖,涵蓋了屬內(nèi)全部3個(gè)染色體(tǐ)組型。栽培種基因組拼接大小(xiǎo)為(wèi)312.95 Mb,6個(gè)野生(shēng)種基因組拼接大小(xiǎo)為(wèi)300.7 Mb(S. angustifolium)- 668.4 Mb(S. radiatum)。BUSCO結果顯示,7個(gè)基因組的(de)拼接完整度為(wèi)91.2-98.5%。
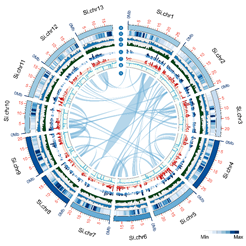
圖1. 芝麻基因組圈圖
基因組直系同源基因Ks分布結果顯示,這(zhè)7個(gè)基因組均經曆了全基因組複制(WGD)和(hé)基因組三倍體(tǐ)化(huà)(WGT-γ)事(shì)件(jiàn)。此外(wài),S. radiatum 還(hái)發生(shēng)有(yǒu)第二次全基因組複制(WGD-2)事(shì)件(jiàn),表明(míng)該種為(wèi)四倍體(tǐ)(圖2A)。7個(gè)種的(de)染色體(tǐ)組原位雜(zá)交(GISH)及着絲粒重複序列熒光(guāng)原位雜(zá)交(FISH)的(de)結果進一步證明(míng),S. radiatum 基因組包含了兩套不同的(de)着絲粒重複序列(Centromere repeats),為(wèi)異源四倍體(tǐ),被命名為(wèi)AABB(圖2B)。其中,AASra亞基因組與n = 16的(de)二倍體(tǐ)野生(shēng)種 S. latifolium 同為(wèi)AA類,BBSra亞基因組與n = 16的(de)S. angolense、S. calycinum、S. angustifolium等3個(gè)二倍體(tǐ)野生(shēng)種為(wèi)BB類。二倍體(tǐ)栽培種 S. indicum (n = 13)和(hé)S. alatum (n = 13)具有(yǒu)各自(zì)特異的(de)着絲粒重複序列,分别被命名為(wèi)CC和(hé)DD類基因組(圖2C)。
單拷貝同源基因進化(huà)分析顯示,胡麻屬分化(huà)發生(shēng)在第三紀時(shí)期的(de)48.5–19.7百萬年(nián)前(MYA)(圖2C)。其中n = 13的(de)野生(shēng)種S. alatum(DD)最早先分化(huà)出來(lái),随後是栽培種(n = 13, CC),其次是n = 16的(de)野生(shēng)種S. latifolium(AA)。S. latifolium分化(huà)時(shí)間(jiān)在34.2 MYA,其他(tā)四個(gè)野生(shēng)種 (n = 16) 的(de)分化(huà)時(shí)間(jiān)為(wèi) 27.2–19.7 MYA。推測S. radiatum異源四倍體(tǐ)化(huà)的(de)時(shí)間(jiān)應該在~5 MYA。在該時(shí)期有(yǒu)多種植物(wù)發生(shēng)多倍體(tǐ)化(huà)事(shì)件(jiàn)。研究認為(wèi)胡麻屬物(wù)種的(de)進化(huà)路(lù)線為(wèi):n = x= 13→ n= x= 16→ n = 2x = 32;物(wù)種向着染色體(tǐ)數目增多的(de)方向進化(huà)。胡麻屬不同種的(de)長(cháng)時(shí)期進化(huà)特點,也反映出該屬物(wù)種在雙子(zǐ)葉植物(wù)進化(huà)中的(de)相(xiàng)對(duì)古老(lǎo)地(dì)位。
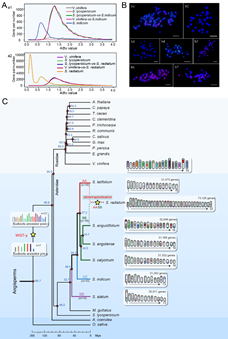
圖2. 胡麻屬基因組進化(huà)特征
泛基因組分析結果顯示,胡麻屬7個(gè)基因組共有(yǒu)25,401個(gè)基因家(jiā)族,包含202,685個(gè)putative基因(圖3)。核心基因家(jiā)族(Core
families)共有(yǒu)9,164(36.1%)個(gè);保守線性基因家(jiā)族比例較大豆屬(Glycine,50.1%)和(hé)棉屬(Gossypium,61.8%)的(de)低(dī),進一步表明(míng)胡麻屬物(wù)種的(de)發生(shēng)應處于重要的(de)物(wù)種擴展時(shí)期。
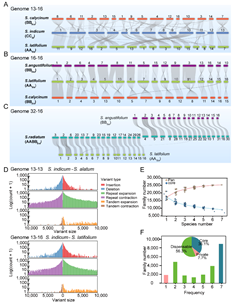
圖3. 胡麻屬7個(gè)種基因組共線性比較與泛基因組分析
同時(shí),研究小(xiǎo)組系統分析了栽培種及野生(shēng)種的(de)基因組結構變異(Structural variations,SV)特征,發現(xiàn)栽培種基因組中發生(shēng)有(yǒu)多個(gè)SV變異,并對(duì)芝麻關鍵農(nóng)藝性狀産生(shēng)重大影響。例如,4166bp的(de)Copia_LTR_反轉座子(zǐ)序列插入和(hé)24.9 Kb的(de)大片段基因組序列缺失,分别引發了PEBP家(jiā)族中的(de)SiPT1和(hé)SiDT1基因發生(shēng)SV變異,進而導緻芝麻株型和(hé)花(huā)序類型性狀發生(shēng)改變(圖4)。
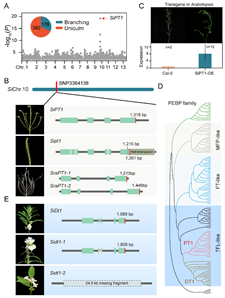
圖4. 胡麻屬株型和(hé)花(huā)序類型調控基因
芝麻枯萎病是芝麻兩大真菌病害之一。與已測序的(de)S. calycinum、S. latifolium、S. angolense、S. radiatum 等高(gāo)抗病野生(shēng)種相(xiàng)比,芝麻栽培種對(duì)強緻病力枯萎病菌F. oxysporum
f. sp. sesami (FOS)菌株則表現(xiàn)為(wèi)高(gāo)感水(shuǐ)平。為(wèi)揭示栽培種與野生(shēng)種在枯萎病抗性方面的(de)差異,研究小(xiǎo)組開(kāi)展了S. calycinum(高(gāo)抗枯萎病)×S. angustifolium(高(gāo)感枯萎病)種間(jiān)雜(zá)交群體(tǐ)的(de)GWAS關聯分析和(hé)SV分析(圖5),成功克隆出芝麻抗枯萎病基因ScaDIR40。該基因編碼Dirigent(DIR)蛋白,調控木酚素和(hé)木質素合成過程中的(de)有(yǒu)機(jī)松柏醇類耦合反應,并能在次生(shēng)細胞壁發生(shēng)過程中提高(gāo)植物(wù)的(de)抗病抗逆性。在高(gāo)感病野生(shēng)種S. angustifolium中,等位基因Saudir32的(de)5′-UTR -210bp處,插入了一段1411bp
的(de)Copia_LTR_反轉座子(zǐ)(圖
5D)。在高(gāo)感病栽培種中,同源基因Sindir37
的(de)-1bp到-1400bp的(de)5′-UTR 序列以及部分CDS序列則發生(shēng)了丢失(圖5E)。上(shàng)述SVs的(de)獨立發生(shēng)均導緻了DIR基因功能發生(shēng)改變,并最終導緻植株枯萎病抗性喪失。研究為(wèi)芝麻遠(yuǎn)緣雜(zá)交、基因編輯等抗病育種研究提供了重要理(lǐ)論和(hé)技(jì)術支持。
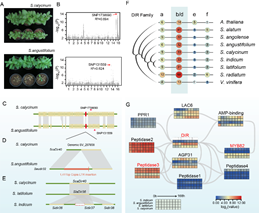
圖5. 胡麻屬枯萎病抗性調控基因DIR
為(wèi)揭示芝麻含油量高(gāo)的(de)分子(zǐ)機(jī)理(lǐ),研究小(xiǎo)組開(kāi)展了油脂合成代謝(xiè)相(xiàng)關基因家(jiā)族(FA)分析(圖6)和(hé)560份芝麻核心種質3年(nián)3點8個(gè)環境下(xià)的(de)GWAS分析(圖7)。發掘出了芝麻高(gāo)含油量基因SiNAC1,首次證實SiNAC1參與調控植物(wù)油脂合成過程(圖7);該基因的(de)T333G堿基突變對(duì)芝麻含油量表型變異的(de)最高(gāo)解釋率為(wèi)43.8%,籽粒含油量可提高(gāo)6.2個(gè)百分點。研究發現(xiàn)NAC1基因在野生(shēng)種中存在結構變異;SiNAC1能夠共表達調控FAD6、NPC1、CYP96A10、FAB1B、FAR2、DPL1、PDAT等不飽和(hé)脂肪酸生(shēng)物(wù)合成與代謝(xiè)調控相(xiàng)關基因(圖 7)。同時(shí),研究小(xiǎo)組還(hái)對(duì)另一個(gè)含油量相(xiàng)關基因SiPPO進行了家(jiā)族分析和(hé)過表達轉基因功能驗證。研究為(wèi)芝麻及其他(tā)油料作(zuò)物(wù)品種改良提供了重要組學信息。
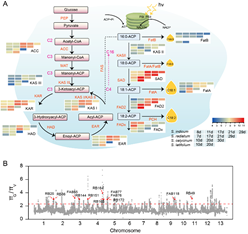
圖6. 芝麻脂肪酸合成路(lù)徑及人(rén)工(gōng)選擇區(qū)域的(de)重要優異基因群分布
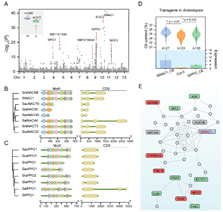
圖7. 芝麻含油量性狀相(xiàng)關基因功能分析
本研究得到了國(guó)家(jiā)特色油料産業(yè)技(jì)術研發中心和(hé)河南省特色油料作(zuò)物(wù)基因組學重點實驗室的(de)支持。

 豫公網安備 41010502004670号
豫公網安備 41010502004670号